���������£����������������������������ǿ�ѧ�������ƶ�������������ѧ���������ˡ�������༭������ͨ���˹�����ø�鵼�Ļ����鶨�����μ��������ּ���ԭ���������κ����ֻ�������κ�λ���Ͻ�������г����Ӷ�������Դ�������������������ġ��˹�����ø�鵼�Ļ�����༭��genome editing with engineered nucleases��������ѡ2011���Nature Methods���ܹ�ע�ļ����ɹ���2013���Science��Nature Biotechnology����־�ϣ������������ɳɴؼ���̻����ظ�����/���ɳɴؼ���̻����ظ����й������ף�clustered regularly interspaced palindromic repeat/CRISPR-associated protein, CRISPR/Cas��ϵͳ��Ϊ������༭��һ����ͨ�ù��ߡ���һϵ�л�����༭�¼������о������ã���һ������������������߳���ʹ�ö�������ó��������ø�Ϊ����Ч����������֡���ϸ������ֻ����Ŵ���������������δ�������ڵõ�Ѹ�ͷ�չ��
ʲô��CRISPR/CASϵͳ��
CRISPR��ָ���ɳɴؼ���̻����ظ����У�clustered regularly interspaced short palindromic repeat����Cas��CRISPR��ص��ף�CRISPR-associated protein����CRISPR/Cas9������༭ϵͳ����ϸ����ϸ���д��ڵĢ���CRISPR/Cas���������ϵͳ���˹�������ɡ���ϵͳ�鵼�Ļ�����༭����Ҳ����RNAָ���ĺ�������ø��RNA-guided endonuclease, RGEN��ϵͳ����ZFN��TALEN������ȣ��ü�������ơ��ϳ���ɸѡ�ϸ�Ϊ��㡢��ݣ����м����ʱ��ͳɱ����ƣ���ͬ��ZFN��TALEN�������ǣ�CRISPR/Cas9������༭ϵͳ������һ��ϸ����ͬʱ���ж������ı༭����˴������˶Ի�����ı༭Ч�ʡ�
CRISPR��������һ��ǰ������leader��������߶ȱ����ظ����У�repeat���ͱ˴���ȫ����ͬ�ļ�����У�spacer����ɣ���������ǰ����������֣����ߴ������������CRISPR���У�ͼ1����ǰ��������ͨ����300~500bp������AT�������ϸ�������ڸ����зdz����أ������ּ�ȴ�����������ظ����еij���һ����23~50bp��ƽ������ԼΪ31bp���ظ�������ͬһ��CRISPRλ�����Ǹ߶ȱ��صģ�һ��ֻ����1~3������IJ��졣���������ּ䣬����ͬһ������Ļ������ϲ�ͬλ�õ�CRISPRλ��֮�䣬�ظ����еı�����ȴ�Ƿdz���ģ����в���ܴ�ͨ����Ŀǰ��֪��CRISPR���������е��ظ����н��з������֣��ظ����а������Ľṹ�����ת¼�����γɷ������Ķ����ṹ���dz��ȶ����ֲ����ظ�����֮��ļ������һ����17~84bp��ɣ�ƽ��������36bp���ҡ�������еı����Էdz��������ͬһ��CRISPRλ���У�Ҳ������û����ͬ�ļ�����С��ظ����еĸ߶ȱ��أ�������е���ȫ��һ�£�ʵ��������CRISPR���е�����ܸ߶���صġ�

ͼ1��CRISPRλ��Ľṹͼ
Cas���Ļ���һ�������λ��CRISPRλ�����Σ�������ʱҲ���ɢ�ֲ��ڻ������С�Cas������ʵ��CRISPR���ܵ���Ҫִ���ߣ���һ���ϴ�Ķ�̬�Լ��嵰�ס�Ŀǰ��Cas���ķ��ಢ��ͳһ������һ���Ǹ���cas�������еı��س̶ȣ���Ϊ�����ͺ���cas��������������cas������ظ��������δ֪���ף�repeat-associated mysterious protein, RAMP������������ࡣ
�����������͵�cas�����У������ͺ���cas����ĵ����ʹ����Ѿ������õ���֤�����磬Cas1��Cas2������Ҫ�������µļ�����жΡ�Cas3��������н���ø�ͺ���ø�Ĺ��ܣ������Ŀ�Ļ�����м��С�����������Cas�����ظ��������δ֪���Ĺ���Ŀǰ�в���ȷ��ֻ��һС���ֵ����ʹ�����֪�����⣬�ܶ�Cas���ײ����ǵ������õģ�������ϳɸ��ӵĵ����������������ã�ͼ2����

ͼ2�����͡����͡�����CRISPR/Casϵͳ�е�Cas���� ������Bhaya et al., 2011��
�ֱ�Դ�ڴ˾���Escherichia coli���������������Streptococcus thermophilus�������ȹ�ϸ����Pyrococcus furiosus����������CRISPR/Casϵͳ�й��е�cas1��cas2�����Ϊ��ɫ��ÿ�����Ͷ��еĵ����ʣ����ͣ�cas3�����ͣ�cas9�����ͣ�cas10����Ϊ��ɫ����ɫ��ǵĵ�����������������Cas��������cas4��cas5��cas6��cas7��
CRISPR/CASϵͳ����Щ���ܣ�
CRISPR/Casϵͳ��ִ�й��ܵķ��֣�ʵ���ϵ����ڲ�������������Ϣ�ķ�չ������������ϸ������������Ϣ�Ļ��ۡ�CRISPR����1987��ͱ����֣���һֱ��2005������о��Ŷ��ƲⲢ֤������������ѧ���ܡ�
CRISPR/Casϵͳʵ�����ǹ�ϸ����ϸ���ڳ��ڽ����������γɵ�����������ϵͳ���ܹ��������������RNAi�ķ�ʽΪϸ���ṩ���߱����������Ե���ֹ���ɾ����Ⱦ�������ӺϺ�ת������ɵĻ�����룬���Ҳ����ΪCRISPR���š�CRISPR/Casϵͳ�����ù�����Ҫ����3���Σ���Ӧ��������š�
�Եֿ��ɾ����ȾΪ����3�������£�ͼ3����
��Ӧ——�¼�����еĻ��
���ɾ���DNA����������д���CRISPRϵͳ��ϸ����ʱ�����������ڵ�CRISPR��ص��������Ѹ������ԴDNA��ϣ�Ȼ��ͨ��Cas1��Cas2�ȵ����ʵ����ã�������ԴDNA�и�ɳ�����17~48bp���ȵ�СƬ�Σ�Ȼ������ص��������£������е�һ��СƬ��������CRISPRǰ�������һ���ظ�����֮�䣬�γ�һ���µļ�����У�ͼ3�������������ͬԴ�IJ���DNA���оͽ���ԭ�����proto-spacer����
ÿһ�β������������ظ����еĸ��ƣ������γ�һ���µ��ظ�-�����Ԫ��������ʹ�ø�ϸ���е�CRISPRλ���б����˴����ɾ����������Ϣ��Ϊ��Ӧ�����ߵ춨�˽ṹ�����������о����֣�CRISPRϵͳ����������ɾ���ĵֿ�����CRISPRλ���ϼ�����еĸ�����ء�������еĸ���Խ�࣬�������ĵֿ�����Խǿ��
����——���ﲢ�ӹ�crRNA
CRISPRλ���У������������������Ϣ�ܱ������������ض��ɾ���Ĺ�����Kunin�ȵ��о�����������CRISPR������λ��ǰ������ĩ�˵�����������ת¼��ת¼�ɰ�������ظ����кͼ�����е�crRNA��CRISPR RNA��ǰ�壨pre-crRNA����֮���ں��ĵ���Cas1-Cas4������ɵĵ�������������£�pre-crRNA��������λ�㱻���п�����ɸ�С��crRNA���������crRNA����Ҫע����ǣ������п���������λ�㱻��Ϊ��λ�ڼ�����еĵ�8�����������Ҳ���о���Ϊ������λ�������ظ������ϡ�
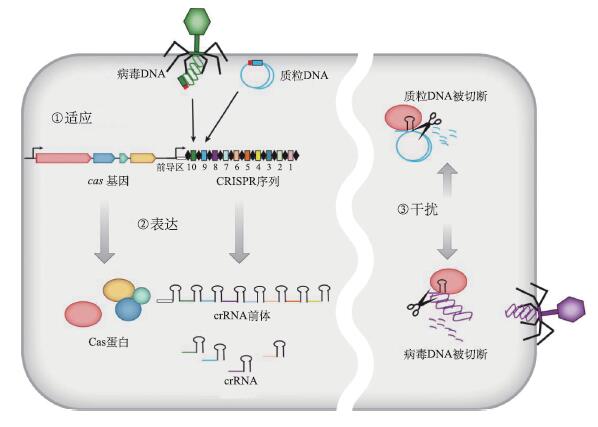
ͼ����CRISPR/Casϵͳ���û��ƣ�����Bhaya et al., 2011��
��һ�Σ��¼�����еĻ�á�Դ���ɾ������������˫��DNA�����ϵ�����CRISPR���е�ǰ������֮��CRISPR���а���������صġ����ظ����зָ��ļ�����У�ͼ�б����ֵIJ�ɫ������Խ�����������ȥ��ʱ��Խ�̣���������еĻ��������ҪCas1��Cas2���İ������ڶ��Σ����ﲢ�ӹ�crRNA��pre-CRISPR RNA��pre-crRNA����RNA�ۺ�øת¼����������Cas���ļӹ����и��СcrRNA��ͼ�еķ��нṹ����ɫ���ִ���������У���ÿ��crRNA����һ�������ļ�����кͲ����ظ����С������Σ�crRNA�ƻ����ֺ��ᡣcrRNA�����ļ���������������ԴDNA���ɾ��������������Ի�����������Cas�����е��иӦ
���crRNA��Cas������������ã���ϳ�һ����������ܵĸ�����crRNP��crRNA-Cas ribonucleoprotein��������һ�η������ã�ͼ3����
����-crRNA�ƻ����ֺ���
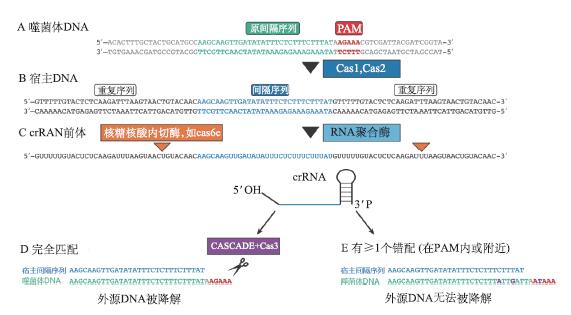
��Ϻõ�crRNP��������crRNA��ƥ�����ã�����������Ѱ�����以�����ɾ���DNAƬ�Σ������������Խ�ϡ��˺����巢�����ã����ɾ���DNA˫�����̣������併�⣬�Ӷ��ﵽ�����Ե���ֹ�ɾ����Ⱦ��Ŀ�ģ�ͼ3������Ҫ�ر�ָ�����ǣ���ʶ����ԴDNAʱ��ԭ������и�����һ�α���Ϊԭ������ڻ���proto-spacer adjacent motif, PAM���Ķ����У��dz����أ�ͼ4��������crRNA����Դ�����ʶ���з�������Ҫ���ã�Ҳ������CRISPR/Casϵͳ���л�����༭�������صĹ���PAM�������ձ�ģ���һЩ�����CRISPRϵͳ�����иû���
ͼ4��CRISPR/Casϵͳ��ԭ������С�������м�ԭ������ڻ�����Bhaya et al., 2011��
A. ����˫��DNA����ɫ���֣���ԭ������У���ɫ���֣���PAM������ɫ���֣��� B. �²�����������ɫ���֣�������DNA��C. RNA�ۺ�øת¼��pre-crRNA��ת¼��ʼλ��δ��ʾ�����Լ�������crRNA����ɫλ��Ϊ����ʱ�ļӹ�λ�㡣D. crRNA����ԴDNA��ȫƥ��ʱ����ԭ����ڲ������и��һ�����У���ҪCASCADE �����弰Cas3���ס� E. ��crRNA����ԴDNA����ȫƥ��ʱ���и������
Ŀǰ�㷺ʹ�õ�CRISPR/Cas������༭ϵͳ�����϶��Ǣ���CRISPRϵͳ����Ϊ����Ģ���CRISPRϵͳ�У�����4��������ɵĻ���أ��ֱ���cas9��cas1��cas2��csn2�����������tracrRNA �����������к��ظ�����������CRISPR���С�����CRISPRϵͳ����Դ˫��DNA���ж����и�Ĺ��̷�Ϊ���¼�����ͼ5����
ͼ5������CRISPRϵͳ�鵼��DNA˫�����ѣ�Cong et al., 2013��
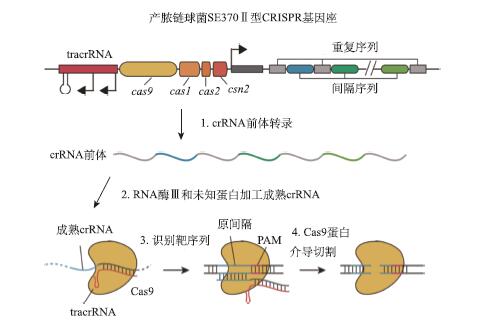
1��CRISPRϵͳת¼��pre-crRNA��tracrRNA��
2��tracrRNA���ݼ���������ԭ����pre-crRNA�γɶ����壬����ص��������£�pre-crRNA���ӹ�Ϊ�����crRNA��
3�������crRNA-tracrRNA������ָ��Cas9������Դ�����еİ����н���ʶ��ʶ�������ͨ��crRNA�ϵļ����������ԴDNA�ϵ�ԭ������еĻ�����ԣ��Լ�PAM���ĸ������ʵ�ֵġ�
4��Cas9�����е�DNA���нṹ������Դ����̶���λ���п�DNA˫����
����CRISPRϵͳ��������Jinek�ȿ�ʼ���죬���ǽ�crRNA-tracrRNA˫��RNA���������ɵ���Ƕ���壬���Ҹ����ĵ���Ƕ�����ܹ�������˫����������ͬ�����ã�ͼ6���������˹�����ĵ���RNA������Ϊָ��RNA��guide RNA, gRNA������һ����ij��֣�Ϊ�˹�����CRISPR/Cas9ϵͳ��ʹ������л�����༭�����˻����������ڸ��о��У����ǻ����֢���CRISPRϵͳ�У�Cas9���װ�����HNH����ø�ṹ�����и���ԴDNA�������л�����������RuvC�ṹ�����и���ԴDNA����һ������

ͼ6������CRISPRϵͳ������Jinek et al., 2012��
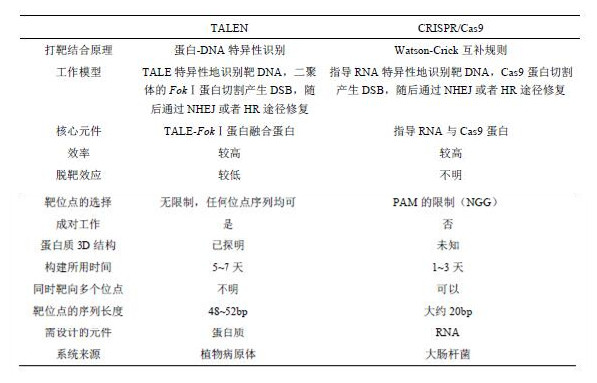
�˺�CRISPR�ڻ�����༭�����д������֣��ں̵ܶ�ʱ���ڣ�����о��ŶӶ��ɹ��ؽ�CRISPR /Cas9ϵͳӦ���������ϸ���еĻ�����༭�У���1������ZFNϵͳ��TALENϵͳ��ȣ�CRISPR /Cas9ϵͳ���ڸ��ָ��ӳ̶ȵĻ����鶼���и��ߵ��������������⣬CRISPR /Cas9ϵͳ�Ĺ�����Ϊ������Cas9�����Է���ؽ�����ø����Ϊ�п�ø��nickase����ֻ��Ҫ��Cas9����������һ����������ͻ�䣨D10A���������и���Ĺ��ܾͱ�Ϊ�и��DNA���ܹ�����ȷ�ؿ���CRISPR /Cas9ϵͳ�Ĵ��Ч��������Ѱеĸ��ʡ��ۺ����������棬CRISPR /Cas9ϵͳ�����ǻ�����༭�������������Ĺ��ߡ�
��1��CRISPR/Cas9������༭ϵͳ�ڻ�����༭�����Ӧ��

CRISPR/CAS������ϸ�������Ӧ��
����CRISPR/Cas9������༭�������м�㡢��ݵ��ص㣬��ѧ����Ҳ���������ڼ���Ӧ���ϵļ���DZ�����ں̵ܶ�ʱ���ڣ�CRISPR/Cas9ϵͳ���ڶ�����ֵĻ�����༭�����������صȷ���õ��˴�����Ӧ�ã���1����2����
CRISPR/Cas9������༭����������ϸ���е�Ӧ��Ч���ѱ�����о�С��֤ʵ����1������Щ�о��漰���ֲ�ͬ����ϸ����������ϸ�����յ��Զ��ܸ�ϸ�����������ͬ����λ�㣨�����Ѿ�ͨ��ZFN����TALEN�ɹ��������εĻ���λ�㣬��CCR5��AAVS1�ȣ����������η�ʽ�����������ó���ͬԴ���顢�������ϡ������ͬʱ�ó��ȣ������˵���˸ü���������ϸ��������༭�е�ǿ�����á�
��ʡ����ѧԺ��Cong����������CRISPR/Cas9������༭����������ϸ����ʵ����EMX1��PVALB˫�����ͬʱ�ó�������EMX1������ó�Ч��Ϊ27%��PVALB������ó�Ч��Ϊ7.3%�����⣬ͨ��ͬʱ�������ͬһ�������������ͬλ���gRNA��ʵ���˶�EMX1����ij���Ϊ196bp��Ƭ��ɾ����ɾ��Ч��Ϊ1.6%������ҽѧԺ��Mali����������յ��Զ��ܸ�ϸ����ʵ����CRISPR/Cas9ϵͳ�鵼�Ļ�����༭�������ǵ��о��У�CRISPR/Cas9ϵͳ�����յ��Զ��ܸ�ϸ���ж�AAVS1λ��Ĵ��Ч��Ϊ2%~4%��
��ࣺ����֪ʶ��